Thank you GWCT
A few days ago we thanked BASC for their response to the Defra consultation on general licences because it clarified how ridiculous was their call for a reversion to the General Licences GL04, GL05 and GL06 which were revoked after Wild Justice’s successful legal challenge.
Today we must thank the Game and Wildlife Conservation Trust (GWCT) for their submission which also appears to favour a reinstatement of the three discredited general licences although not in any way as positive or strident a way as that of BASC. In fact, it is somewhat difficult to find a recommendation for action in the GWCT response.
Where GWCT say in their Summary ‘We highlight scientific evidence that predation control carried out under General Licences can lead to annual increases in breeding densities of a range of red-listed birds (e.g. grey partridges 35% increase per year, curlew 14% increase per year). We also highlight numerous case studies where farmland birds have responded positively to predation control.‘ they fail to disclose that the predation control to which they refer usually involved control of mammals as well as birds and rarely was able to distinguish between impacts of different bird species. The studies cited by GWCT, taken with other important studies which GWCT have neglected to cite, form no basis for any call to reintroduce a licensing regime for bird killing which allows the lethal control of Jay, Jackdaw, Rook or Magpie – the science for an impact of these corvid species on wild bird population levels is weak or non-existent.
What follows is an evaluation of the GWCT argument, carried out by Wild Justice with the help of other scientists, and an assessment of the evidence against four corvid species.
- An assessment and evaluation of the GWCT evidence:
GWCT present a number of lines of evidence to propose that predator removal generally can improve breeding success and sometimes the population sizes of other species, but there is little evidence that deals with corvids explicitly and almost nothing on the impacts of individual species of corvid. Furthermore, GWCT give no consideration to other predator reduction methods. One of the conclusions of a recent major review of predation impacts (Roos et al. 2018), cited by GWCT, was that “Much more attention needs to be paid in future to landscape-scale measures to reduce predation impacts, such as changes in land-use configurations (e.g. removal of commercial forest plantations…), habitat management … and the re-establishment of populations of apex predators …, as well as a critical test of the environmental sustainability of intensive gamebird releasing that may cause trophic cascades…” [references replaced with ellipses]. A further general comment is that predator reduction or removal experiments of the kind that form the crux of GWCT’s evidence are often inherently biased by being undertaken largely in areas where it is supposed that there will be an impact; hence it is difficult to generalise their findings to other situations.
We address GWCT’s arguments in turn here.
- GWCT cite as a “key reference” the study of Macdonald & Bolton (2008) [Predation on wader nests in Europe. Ibis 150: 54-73]. This exhaustive review of a large number of studies from across Europe documents around 3,000 nest predation events recorded by a variety of methods. The majority of predation events could not be assigned to individual predators, and were frequently recorded as nocturnal vs diurnal, or avian vs mammalian. Of those that could be assigned to individual species or species groups, corvids (specifically or generically) were implicated in just 38 (Magpie 1, “crow” 16, “corvid” 21). Of 200 nest predation events, predominantly in wet grassland, identified using nest cameras in which the identity of the predator could be established, 132 (66%) were by Foxes, 21 (10.5%) by Stoats, 15 (7.5%) by Crows, 12 (6%) by Badgers, 8 (4%) by Hedgehogs, and small numbers (2%) by other predators. The study concluded that “The use of temperature loggers initially, and nest cameras more recently, has shown that the widely held belief that birds (particularly corvids) are the major predators of wader nests is frequently not true.”
- GWCT cite their Salisbury Plain and Otterburn predator removal experiments in defence of their stance on the issuing of General Licences, and showed that predator removal led to an increase in breeding success and populations of Grey Partridges (Salisbury Plain) and a number of upland breeding species (Otterburn). However, predator removal at both sites included mammals (Fox, Stoat, Weasel and Rat), as well as corvids (Carrion Crow, Magpie, Jackdaw and Rook). In both experiments, predator control significantly reduced densities of Foxes and Carrion Crows, but there was no assessment of the identity of nest predators before, during or after either experiment, so the impacts of corvid removal could not be assessed explicitly.
- Corvid removal studies are cited as evidence that “the ability to apply targeted corvid control at short notice can be beneficial, where breeding hedgerow nesting and probably other songbirds are exposed to breeding corvids”. They cite three lines of evidence:
- “In their recent comprehensive review Roos et al. (2018) state (in the abstract) that they found little evidence that predation limits populations of passerines but that they [sic] do limit waders. This, however, is not a full and balanced reflection of the results, and a key finding of the review, highly relevant to this call for evidence, is easily overlooked. Table 5 [actually Table 6], which refers specifically to experimental predator removal studies, shows songbirds increased in 40% of 20 studies following predator removal. For waders, it was similar, at 44% of 29 studies. The conclusion from this is that the science available prior to 2017 tells us that corvid removal can lead to an increase in songbird population size.” [our emphasis] We consider this conclusion to be an egregious misinterpretation of the data shown in Tables 5 and 6 of Roos et al., which relate to predator control/removal generally, and not corvid control/removal explicitly. Figure 4 of the same article indicates that the evidence that corvids alone can limit their prey numerically is no different from zero, and Table 7 shows that in only 13.6% of over 360 cases in which the effect of corvids on populations of their prey species have been assessed were the impacts found to be negative (and in 12% it was actually positive).
- The study of Sage & Aebischer 2017 is cited as evidence of a negative impact of corvids on nesting success of hedgerow passerines, although nesting success was measured only indirectly. This study, a rare example of a predator control/removal study that apparently controlled only corvids (though other predator control activities at the sites is not quantified), recorded very modest increases in apparent nesting success of 10-16% across all species with corvid control, but not for open nesters or hole nesters separately and with results that varied between sites – at some sites, nesting success was higher in the absence of corvid removal. The authors concluded that the impacts of this difference in nest productivity on overall population size of songbirds fell somewhere between unlikely and unquantifiable.
- The third piece of evidence is in the form of an unpublished PhD thesis, which we have not seen and therefore upon which we cannot comment, other than to note that one chapter appears to use artificial nests as a surrogate for real nests, a methodology that has come in for severe criticism (e.g. Moore & Robinson 2004, Ecology, 85: 1562-1567).
- GWCT cite their research at Loddington, Arundel and Royston, but this shares the same problems as the Salisbury Plain and Otterburn projects described above, in that predator control was general, including mammals as well as birds, that the identity of nest predators was not known, and that the impact of individual corvid species on populations of other species cannot be evaluated. White et al. (2008) concluded from their study of blackbird nesting success that “Although predator control appeared to have a positive influence on Blackbird breeding population size, the non-experimental set-up meant we could not eliminate other potential explanations”. There is a suggestion in White et al. (2014) (Loddington study) that the contribution of corvid control to the broader patterns noted was negligible: “Sporadic corvid reduction had a positive effect on nest survival only for common blackbird (at the nestling stage only) and a negative effect only for yellowhammer (across both stages)”.
- GWCT describe an unpublished study from the Avon Valley which suggests that unidentified daytime predators (presumed but not proved to be corvids) of Lapwing nests account for 41% of observed predation events or for 41% of all nests (the results are ambiguously presented). They describe a correlation between Lapwing nest survival and Carrion Crow density, but rightly point out earlier in their evidence that “statistically significant correlations do not indicate cause and effect and can be caused by unmeasured factors”. We have not seen this unpublished work so cannot comment further.
- GWCT describe an unpublished study from across the UK on breeding Curlews, finding that general predator control was associated with higher numbers and breeding success of Curlews. It is suggested that there is a negative correlation (the strength of which is not reported) between Curlew nesting success and Carrion Crow density. This evidence is used to “outline[s] the potential fate of one of the UK’s most threatened bird species if corvids are not legally controlled”, with the conclusion that “expanding predator management, particularly of corvids, currently done routinely on grouse moors, to peripheral unkeepered areas is a practical step that could quickly help stem the current rapid decline of curlew” [our emphasis]. It is not possible to assess from the data presented whether or not the presumption that crows are primarily responsible for the observed differences in numbers and breeding success is justified. However, the results appear to broadly support those of Douglas et al. (2014) [J. Appl. Ecol. 51: 194–203], who concluded that: “direct predator control may also be important to conserve ground-nesting birds in these landscapes, for example, where moorland management and forestry coexist as major land uses”.
- We cannot comment on the Member Responses as the claims made therein are not supported by empirical evidence.
The GWCT consultation response does not cite a number of other relevant studies:
Thomson et al. (1998) The widespread declines of songbirds in rural Britain do not correlate with the spread of their avian predators. Proc. R. Soc. Lond. B 265: 2057-2062. This paper analysed population trends of songbirds from 1965-1995 and assessed the likely impact of Magpies and Sparrowhawks. They concluded that: “magpies and sparrowhawks are unlikely to have caused the songbird declines because patterns of year-to-year population change did not differ between sites with and without these predators”.
Newson et al. (2010) Population change of avian predators and grey squirrels in England: is there evidence for an impact on avian prey populations? J. Appl. Ecol. 47: 244–252. This paper, co-authored by GWCT, concludes that “Analyses of large-scale and extensive national monitoring data provides little underlying evidence for large-scale impacts of widespread avian predators and grey squirrels on avian prey populations, although we cannot exclude the possibility that a small number of negative associations between particular predator and prey species reflect causal relationships or that predators affect prey species at smaller spatial scales.” Writing explicitly of the impacts of corvids, the authors stated that: “For the corvids no significant negative associations were found.”
Madden et al. (2015) A review of the impacts of corvids on bird productivity and abundance. Ibis 157: 1-16. The authors of this review of 42 different studies found that: “Experimental studies that removed only corvid species were significantly less likely to show a positive impact on productivity than those removing corvids alongside other predators (16 vs. 60%). This suggests that the impact of corvids is smaller than that of other predators, or that compensatory predation occurs” and concluded that “These results suggest that in most cases bird populations are unlikely to be limited by corvid predation and that conservation measures may generally be better targeted at other limiting factors. However, negative impacts were found in a minority of cases, and those may require further investigation to develop management tools to mitigate such impacts where they are of economic or conservation concern.”
- Assessing the impacts of individual species: Jackdaw, Rook, Jay and Magpie
It is very difficult or impossible to extrapolate from the many studies listed above the impacts of individual corvid species. In most studies on the effects of predator control, no information is given on how many of each species were controlled and none on the identity of nest predators, and hence it is often hard even to separate out the effects of bird vs mammal predation, let alone the contributions of individual bird species.
Below we summarise the results of three key recent reviews (Macdonald & Bolton 2008, Madden et al. 2015 and Roos et al. 2018 – full references given above) on the potential impact of each species on the productivity and numbers of potential prey species.

Jackdaw:
This species is largely granivorous in winter and largely insectivorous in the breeding season, but in some situations, highly specialised individuals may take the eggs of other birds (Handbook of the Birds of the World).
Macdonald & Bolton (2008) contains no explicit mention of Jackdaw, which was not identified as a wader nest predator in any study. Madden et al. (2015) identify one study of the possible impact of Jackdaw on productivity and four studies of the possible impact of Jackdaw on numbers of potential prey species; in all five cases, no impact was detected. Roos et al. (2018) also do not mention Jackdaw in the list of species whose impact on other species was assessed.
Conclusion: GWCT do not make a case why Jackdaw should be killed under an unregulated open general licence. They do not present evidence that shows any serious impact of Jackdaw on bird populations, and we can find no such evidence. Without such evidence Defra cannot issue a scientifically valid general licence.

Rook:
Main food items include earthworms (Lumbricidae) and grain; a wide range of other small invertebrates also taken, notably beetles (Coleoptera) and cranefly larvae (Tipulidae), also such vertebrates as small lizards, frogs and small mammals, and eggs and nestlings of small birds (Handbook of the Birds of the World).
Macdonald & Bolton (2008) contains only one explicit mention of Rook, from an unpublished doctoral study that suggested that while wader nest predation rates were higher near to rookeries, they were lower on sites with rookeries than on sites without rookeries. Neither Madden et al. (2015) nor Roos et al. (2018) identified any studies of the possible impact of Rook on the productivity or numbers of potential prey species.
Conclusion: GWCT do not make a case why Rook should be killed under an unregulated open general licence. They do not present any evidence that shows any serious impact of Rook on bird populations, nor can we find any. Without such evidence Defra cannot issue a scientifically valid general licence.

Jay:
Chiefly eats invertebrates during breeding season, notably caterpillars and beetles (Coleoptera) gleaned from foliage of trees; diet also includes eggs and nestlings of a range of birds up to size of a Sparrowhawk (Accipiter), much less frequently killing adult passerines, and is among the most frequent predators of other small birds’ nests in some areas; wide variety of seeds and berries eaten, especially in autumn and winter, including grain, beech mast, chestnuts and acorns. (Handbook of the Birds of the World).
Macdonald & Bolton (2008) identified no studies of the possible impact of Jay on the productivity of wader nests, though differences in habitat selection are unlikely to bring Jays into contact with breeding waders. Madden et al. (2015) did not include Jay in their review. Roos et al. (2018) identified 59 cases where the evidence that Jay can limit prey species has been assessed. Of these, 3 (5.1%) indicated a negative impact of Jays, whereas 13 (22%) indicated a positive impact.
Conclusion: GWCT do not make a case why Jay should be killed under an unregulated open general licence. They do not present evidence that shows any serious impact of Jays on bird populations, nor can we find any. Without such evidence Defra cannot issue a scientifically valid general licence.

Magpie:
Omnivorous, but chiefly a carnivorous scavenger. Diet varies according to local habitats, basically comprises invertebrates, especially beetles (Coleoptera), and small mammals and lizards, frogs, bird eggs and nestlings, as well as carrion and rarely even adult birds.
Macdonald & Bolton (2008) identified Magpie as a predator of wader nests in just one study, in which it accounted for a single nest predation event. Madden et al. (2015) identified 41 cases where the impacts of Magpie on the productivity of potential prey species could be assessed, of which 4 showed a positive effect, 5 a negative effect and 32 no effect. Similarly they identified 96 cases where the impacts of Magpie on the numbers of potential prey species could be assessed, of which 6 showed a positive effect, 4 a negative effect and 86 no effect. Roos et al. (2018) identified 104 cases where the evidence that Magpie can limit prey species has been assessed. Of these, 6 (5.8%) indicated a negative impact of Magpies, 16 (15.4%) indicated a positive impact and 82 (78.8%) indicated no effect.
Thomson et al. (1998) concluded that Magpies were not implicated in the decline of songbirds across the UK. Sage & Aebischer (2017) found a small effect of corvid (Carrion Crow and Magpie) removal on the nesting success of hedgerow passerines but could not demonstrate whether this was the result of the removal of Magpies or of Carrion Crows, suggesting that the former was more likely on ecological grounds.
Conclusion: GWCT do not make a case why Magpie should be killed under an unregulated open general licence. They do not present evidence that conclusively demonstrates a serious impact of Magpie on bird populations, nor can we find any. Without such evidence Defra cannot issue a scientifically valid general licence.
- Summary
We consider that the GWCT evidence fails to alter the case against the issue of General Licences for the control of corvids for the following reasons:
- The evidence of the GWCT’s own research does not and cannot, in almost all cases, distinguish between the impacts of corvids and the impacts of other bird and mammal predators. Other scientific literature is in several places misconstrued by GWCT to indicate an impact of corvids whereas the evidence being referred to relates to the impacts of general predator control
- Recent research indicates that mammals are generally much more important predators of birds’ nests than are corvids, but even for mammals there is scant evidence that nest predation influences subsequent population sizes
- The evidence of a number of key recent reviews is not considered in the conclusions reached; these generally conclude that there is no effect of corvids on the populations of other species
- The case against corvids generally is weak, but the case against individual species is either entirely lacking (Rook, Jay, Jackdaw) or almost so (Magpie).
Nowhere in their submission do GWCT actually state that there is sufficient or strong evidence to justify lethal control of individual corvid species under the past or any future licensing system. Their review of the science insinuates that view but does not provide evidence for it.
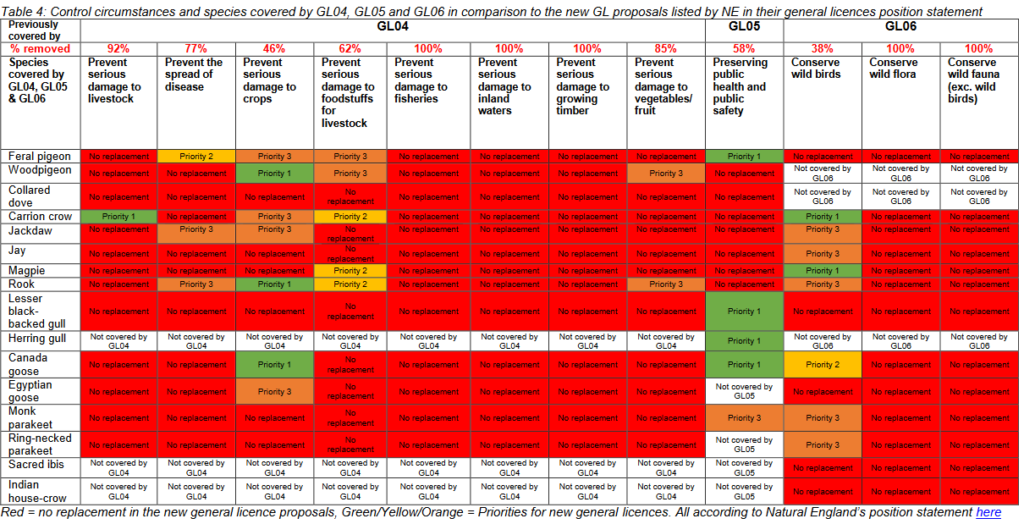
As Defra officials and Michael Gove look carefully at each cell in the Table usefully provided by BASC they must decide, on the basis of the science, whether there is scientific evidence to justify the issuing of licences for lethal control of each species, one by one, for the purpose of conserving wild birds etc. The GWCT response to the consultation will aid them in that it shows how poor is the case for lethal control of many corvid species.
Wild Justice remains steadfast in its call to Defra not to issue future general licences which authorise the lethal control of Jays, Jackdaws, Rooks or Magpies for the purpose of conserving wild birds because the scientific evidence to support such a policy is either weak or, more often, entirely lacking. The days of killing these four species under the pretence that such actions are backed by science or that they are an aid to nature conservation should be left in the past.